裏の裏を行く RNA 配列
 加藤泰彦
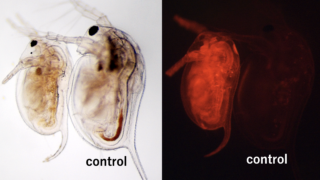
加藤泰彦転写因子をコードするミジンコの Dsx1 遺伝子は胚発生過程でオス特異的に発現し、オス化のマスター因子として働きます。この Dsx1 遺伝子の 5´ UTR 配列 205 塩基が、変幻自在に機能を変えることがこれまでの私達の研究でわかってきました。
その一つとして Dsx1 遺伝子の 5´ UTR としての機能、 Dsx1 mRNA の翻訳抑制が挙げられます。翻訳抑制は 5´ UTR に RNA 結合タンパク質 Shep が結合することで生じます。興味深いことに Shep は雌雄両方で発現することも明らかにしました。これらのことから、 Dsx1 遺伝子の発現が抑制されている雌では、 Shep はノイズによって生じる Dsx1 mRNA の翻訳を抑制することで Dsx1 によるオス化を防いでいると考えられます。雄においても同様に Shep は Dsx1 mRNA の発現の閾値を上げることで、オス化のシグナル伝達のファインチューニングをしていると予想されます。これが、 5´ UTR 配列の表の顔です。
意外なことに Dsx1 遺伝子の 5´ UTR 配列は裏の顔を持つこともわかってきました。Dsx1 遺伝子座からは Dsx1 mRNA のみならず Dsx1 遺伝子を活性化する lncRNA ダパールがオス特異的に発現します。lncRNA ダパールは Dsx1 の転写開始点の上流から Dsx1 mRNA と同じ向きで Dsx1 5´ UTR を含むように転写されます。ダパールの全長を個体内で過剰発現すると Dsx1 mRNA の翻訳が促進されます。Dsx1 5´ UTR のみをコードする RNA を過剰発現させてもダパールと同様に Dsx1 の翻訳促進が見られます。一方で、ダパールから Dsx1 5´ UTR の配列を除くと翻訳促進能が消失します。これらの in vivo 解析に加え in vitro 解析を行った結果、5´ UTR 配列が lncRNA ダパールの中に含まれことで 「細胞質内で」 Shep のデコイとして機能し、Dsx1 遺伝子の発現を脱抑制することを見出しました。
さらに私達は最近、ダパールに含まれる Dsx1 の 5´ UTR 配列が「核内で」 Dsx1 を活性化をしていることを示唆する結果も得ました。本研究では、この裏の裏の顔までをも持つ 5´ UTR 配列においてそれぞれの機能が発揮される時に形成される 2 次構造および 2 次構造に結合するタンパク質を明らかにすることを目指しています。本研究を通して変幻自在に働く RNA 型非ドメインバイオポリマーの作動原理の解明に少しでも貢献できればと考えております。これからどうぞよろしくお願いいたします。

-320x180.png)