非常識にもほどがある!RNA依存性DNA組換え酵素の発見
 西増 弘志
西増 弘志我々は最近、Arc InstituteのPatrick D. Hsu博士らとの共同研究により、ブリッジRNAと協働してDNA組換え反応を触媒する驚きの酵素「IS110リコンビナーゼ」に関する2報の論文を発表しました(文献1、2)。本記事では、IS110システムについて、研究の背景なども含めて紹介したいと思います。
詳細は原著論文、プレスリリース、動画をご覧ください。
プレスリリース:https://www.rcast.u-tokyo.ac.jp/ja/news/release/20240627.html
動画:https://www.youtube.com/watch?v=hJ7zvZTxgT4
挿入配列(IS; insertion sequence)因子は、原核生物のゲノムに存在する転移因子であり、1980年代から研究され、約30のファミリーに分類されています。通常のIS因子はトランスポザーゼをコードしており、このトランスポザーゼがIS因子の両端に存在する特異的な塩基配列(逆向き反復配列)を認識し、ゲノムDNAの別の部位にIS因子を挿入します。一方、2000年代までの研究から、IS110ファミリー因子は通常のIS因子と異なり、(1)長い逆向き反復配列を持たない、(2)環状DNA中間体としてゲノムDNAから切り出される、(3)特定の配列を持つゲノム領域に転移する、(4)(ターゲットDNAに対する特異性が高いことと一致して)通常のトランスポザーゼとは異なるリコンビナーゼをコードする、といった特徴をもつことが明らかになっていました(文献3)。しかし、IS110ファミリー因子の転移メカニズムは20年以上不明のままでした。
そのような状況の中、Patrickらは、IS110ファミリー因子に分類される大腸菌由来IS621因子が、IS621リコンビナーゼに加えて非コードRNA(ブリッジRNAと命名)を産生することを発見しました。ブリッジRNAの発見がこの研究の最大のブレークスルーでした。さらに、大腸菌を用いた実験系において、IS621リコンビナーゼとブリッジRNAが協働し、ブリッジRNAの一部の領域(ガイド配列)と相補的なドナーDNAとターゲットDNAの間の組換え反応を触媒することを見出しました。
2022年11月、Patrickからのメールをきっかけに共同研究が始まりました。IS621リコンビナーゼは大腸菌由来のタンパク質ですが、大腸菌発現系では組換えタンパク質がほとんど得られないということだったので、生化学実験用のタンパク質調製からプロジェクトに参加しました。我々もIS621リコンビナーゼを大腸菌で発現させ精製してみましたが、ほとんど得られませんでした。そこで、助教の平泉くんが、大腸菌発現系の条件検討に加え、昆虫細胞発現系を試してくれました。その結果、昆虫細胞でIS621リコンビナーゼはよく発現し、高純度に精製できることがわかりました。大腸菌由来のタンパク質を昆虫細胞で発現させるという発想は一般的ではありませんが、平泉くんは、以前から膜タンパク質を昆虫細胞や哺乳類細胞で発現させてきた豊富な経験があったため、迅速に昆虫細胞発現系に切り替えることができました。これもこの研究のブレークスルーのひとつでした。
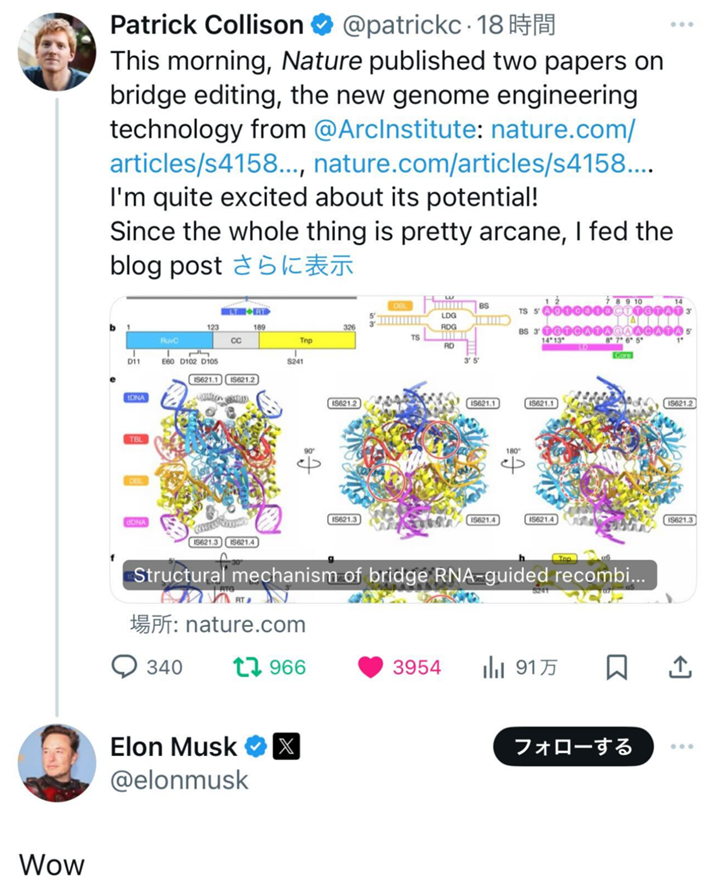
精製酵素を用いた生化学実験の結果、IS621リコンビナーゼがブリッジRNAと複合体を形成し、ブリッジRNAと相補的なドナーDNAおよびターゲットDNAの間の組換えを触媒することが確認されました。さらに、クライオ電子顕微鏡解析により、IS621リコンビナーゼ-ブリッジRNA-ドナーDNA-ターゲットDNA複合体の立体構造を決定しました。構造解析の結果、この複合体はドナーDNAとターゲットDNAの二本鎖の片方(トップ鎖)が切断された状態を捉えたものであることがわかりました。さらに、決定した複合体構造から、IS621リコンビナーゼはCreリコンビナーゼ(チロシンリコンビナーゼ)と配列相同性を持たないにもかかわらず、DNA組換えメカニズムに予想外の類似性を持つことが示唆されました。そこで、トップ鎖交換後の状態が安定化するような配列を持つドナーDNAとターゲットDNAを設計し、クライオ電子顕微鏡解析を行ったところ、予想した通り、トップ鎖交換後の状態の構造決定に成功しました。また、幸運なことに、1種類の複合体試料から、トップ鎖交換後にボトム鎖が切断される前後の2つの状態の構造を決定することができました。これら3つの反応中間体の立体構造から、IS621リコンビナーゼによるブリッジRNA依存性DNA組換えの詳細な分子メカニズムを明らかにすることができました。クライオ電子顕微鏡解析技術がなければ、研究開始から約1年で3つの状態の構造決定に成功し、IS621システムの発見から構造解析までの結果をback-to-backで発表することは不可能でした。
CRISPR-Cas9はガイドRNAと相補的なターゲットDNAを切断する「プログラム可能な」DNA切断酵素であるため、ゲノム編集などさまざまな技術に応用されてきました。ブリッジRNAのガイド配列も変更可能であるため、IS621システムを用いることで、さまざまな配列を持つドナーDNAとターゲットDNAの間の組換えが可能です。したがって、IS621リコンビナーゼは「プログラム可能な」DNA組換え酵素として、CRISPR-Cas9では不可能だった長鎖DNAの組み込みなどの幅広い応用が期待されます。また、IS110ファミリー因子は多様なリコンビナーゼとブリッジRNAをコードしていることがわかっているので、これらの構造機能研究も待たれます。
今回の研究から、IS621リコンビナーゼ-ブリッジRNA複合体はドナーDNAとターゲットDNAを認識し、4本のDNA鎖のうち2本(トップ鎖)を切断・交換・再結合した後、残りの2本(ボトム鎖)を切断・交換するという極めて複雑な反応を触媒することが明らかになりました。自然界には想像を超える酵素が存在することをあらためて認識しました。また、このような驚くべきシステムが最も身近な細菌である大腸菌に存在していたことも驚きです。今回の研究を通じて、人とのつながりの重要性も再認識しました。PatrickはFeng Zhangラボ出身で、2014年に発表したCRISPR-Cas9の構造解析論文の共著者でした(文献4)。それから10年が経ち、互いに研究室を主宰し、新たな共同研究を通じて大きな発見を成し遂げられたことを非常にうれしく思います。IS621因子は2003年に東京大学分子細胞生物学研究所の大坪栄一先生らのグループによって報告されたIS因子です(文献3)。奇遇なことに、私はその当時、同じキャンパスで東京大学大学院農学生命科学研究科の大学院生として研究していました。したがって、どこかで大坪先生とすれ違っていたかもしれません。20年の時を経て、IS621因子の転移メカニズムを明らかにできたことに数奇な運命を感じます。
文献
- Durrant, M. G. & Perry, N. T. et al. Bridge RNAs direct modular and programmable recombination of target and donor DNA. Nature 630, 985–993 (2024).
- Hiraizumi, M. et al. Structural mechanism of bridge RNA-guided recombination. Nature 630, 994–1002 (2024).
- Choi, S., Ohta, S. & Ohtsubo, E. A novel IS element, IS621, of the IS110/IS492 family transposes to a specific site in repetitive extragenic palindromic sequences in Escherichia coli. J. Bacteriol. 185, 4891–4900 (2003).
- Nishimasu, H. et al. Crystal structure of Cas9 in complex with guide RNA and target DNA. Cell 156, 935–949 (2014).
イーロンマスクも注目!
投稿者プロフィール