RNAとタンパク質の液―液相分離によるゲノム制御
 安原 崇哲
安原 崇哲この度、公募班で参加させて頂きます安原崇哲と申します。これまでDNA修復など、ゲノムの安定性を専門としていたのですが、RNAやタンパク質の凝集体などがゲノム安定性を制御するかもしれないということが最近分かってきて、この領域で解き明かそうとしているような様々な現象に非常に興味をもっています。
DNA修復の際には、修復タンパク質がDNA損傷部位に局所的に集合して核内のフォーカスとして観察されるのですが、最初に液―液相分離の概念が提唱され始めたころ、修復分野の多くの研究者は、このフォーカスこそが液―液相分離でできているのだと思いました。確かにそういうことを示唆する論文もいくつかあるにはあるのですが、色々調べてみると、RNA結合系のタンパク質に比べて、DNA修復系のタンパク質は(案外?)しっかりとした構造をとっていて、少なくともDNA修復フォーカス自体は、どうもあまり液―液相分離ぽくはないぞという流れになっています。
そんなころ、RNAポリメラーゼIIの阻害剤が染色体転座のようなゲノム異常を増加させるというデータを得て、転写阻害によっておこるゲノム構造の変化が、2つのDNA損傷部位を近づけて、転座のような異常を引き起こすのではないかと考えました。しかし、転写阻害によって一体何がそんなに変化するのかさっぱりわからず、しばらく停滞していました。基本転写因子の複合体の中に液―液相分離を起こすようなモチーフをもつタンパク質がないか、一つずつ天然変性領域予測ツールに入れてちまちま調べたりしていたのですが、その時見つけたのがTAF6というTFIIDのサブユニットでした。免疫染色ができるTAF6の抗体を買おうとサイトを調べていたら、たまたま似たような名前でTAF15というタンパク質の抗体があって、バリデーションの免疫染色の画像がとても綺麗だったので何となく一緒にカートにいれて注文していました。
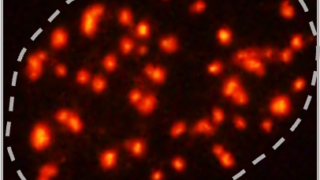
結局、TAF6を免疫染色しても、GFPをつけて発現させても、転写阻害で局在は微動だにせず、そもそものアイデア自体をあきらめかけていたのですが、そういえばTAF15の抗体せっかく買ったのに使ってないし、最後に免疫染色でもみておくかと思って、何も期待せずにスライドを見てみたところ、美しい光景が広がっていました…。
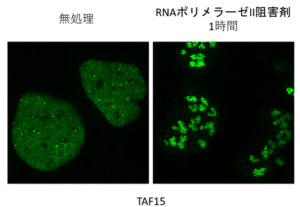
その時はまだ知らなかったのですが、このTAF15というタンパク質、TBP-Associated Factor 15でいかにも基本転写因子のサブユニットぽい名前をしているのですが、構造的にはむしろFUSとかEWSなどのRNA結合タンパク質のファミリーに属していて、文献をみると明らかに液―液相分離を起こすタンパク質でした。
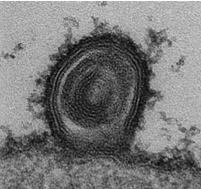
そこからは芋づる式にSFPQ、NONO、FUSなどのパラスペックル因子がこの核小体に形成される凝集体を構成しており、足場となるRNAに関しては、NEAT1ではなく、未プロセシング状態のリボソームRNAが必要ということがわかってきました。この凝集体が本当にゲノム構造を変化させるのかというところは一苦労したところでしたが、TSA-seqとよばれる局所的ビオチン化法によるシークエンス解析を行うことで無事証明することができ、リボソームRNAを足場とした凝集体の形成がゲノム構造を変化させ、それによって染色体転座が発生するというストーリーを描くことができました。
当領域では引き続きこの核小体に形成される機能不明な凝集体の性状を明らかにしながら、ゲノム構造を意図したようにコントロールするようなツールとして使うことができないか検討しようと思っていますので、ぜひ領域の皆様から色々な視点でご指導いただければありがたいです。