#20の大迷走(2)
 NakagawaShinichi
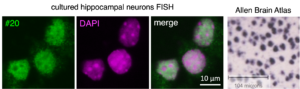
NakagawaShinichi6330403K07Rikの核内局在はadult mouse brainで最初確認していたのですが、海馬神経細胞の初代培養で見てみると、培養初期にはほとんど発現しておらず、強いて言えば細胞質にシグナルが若干発現が見られます。神経細胞の成熟に従って発現が上昇し、核内に強いシグナルが見えるようになります。Neruo2A細胞でも、核に強いシグナルが観察されました。
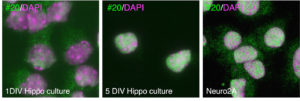
海馬培養細胞では成熟に伴って#20のシグナルは強くなり、N2Aでも核内にシグナルが見える、というのが幻だったとはこのときは知るすべもなく、、、
というわけで、核に局在する機能性lncRNA候補6330403K07Rikが見つかって小躍りしていたわけですが、当初のアノテーションではlncRNAとされていたこの遺伝子。よくよくその配列を見ると、結構しっかりとしたORFがあるではありませんか。そしてそのORFはヒトでも保存されています。うーむ。これはやはりlncRNAではなくて何らかのタンパク質をコードする遺伝子なのか。しかもC'側には疎水性のストレッチがあって、シグナルペプチドはなく、ゴルジかER関連のタンパク質っぽい感じです。

よくよく見ると保存されたORFが、、、核内lncRNAの研究から核内繋留mRNAの研究になったところが引き返しどころだった?
ここが、最初の、引き返すべきポイントだったのかもしれません。
とはいえ、スプライシングされたmRNAがここまできれいに核に局在しているのであれば、それはそれで珍しい。ただ、6330403K07Rikは2 exons geneで、そのORFはタンパク質をコードするmRNAとしては珍しく1st exonに入っているので、NMDの標的となって細胞質に輸送されたmRNAがNMDで分解されただけということも考えられます。でも、cyclohexamideで翻訳を止めても細胞質のシグナルが見えてくる気配はありませんでした。これは何らかの未知のメカニズムでmRNAが核内に繋留されているはず!面白い!と、己の変わり身の速さに呆れつつ、敬愛する先輩の偉大な名言「手のひらは何回ひっくり返しても減らんからな」に則って、本プロジェクトの看板を、lncRNA研究から、mRNAの核内繋留メカニズム研究にかけかえることにしました。
核内繋留されるのであれば、何らかの繋留シグナルがmRNA内にあるはずです。5' UTRなのか?3' UTRなのか?はたまたORFなのか?とりあえずそれぞれの領域を切ったり貼ったりした変異mRNAをHeLa細胞に発現してみたところ、全長のmRNAであれ、deletionのRNAであれ、ことごとく細胞質に運ばれていきます。
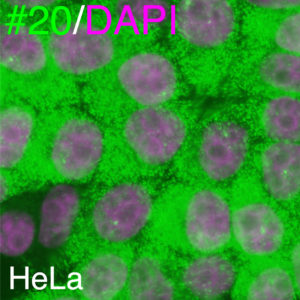
核内繋留シグナルを同定しようと全長、欠失変異体、イントロンありなし、ありとあらゆる組み合わせを試しましたがことごとく細胞質に運ばれてしまう#20のmRNA
このあたりで立ち止まってじっくり考えるべきだったのでしょうが、ハマるときはハマるもので、むしろ6330403K07RikのmRNAを核内繋留するトランスの因子がHeLaにはないに違いない!神経細胞に導入すれば核局在するかも!
しかしながら、悲しいかな。海馬ニューロンの初代培養に導入しても細胞質に運ばれたのでこの仮説はボツ。
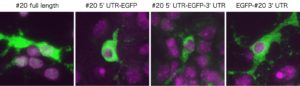
海馬初代培養ニューロンに導入しても、やはり細胞質に運ばれてしまう#20とその変異分子ののmRNAたち。核内繋留シグナルは一体どこに??
であれば、matureなmRNAではなくてpre-mRNAがスプライシングされる際に核内繋留因子がロードされるに違いない!とこれはもう謎が解けた、もしそうならプチ大発見だ、と意気込んだものの、イントロン付きのpre-mRNAを発現させても細胞質に運ばれるのでこの仮説もボツ。
も、もしかして6330403K07Rikのプロモーターから転写されたmRNAは核内に繋留されるなんて大発見??と、これはもう思いついたときは興奮して寝られないぐらいはしゃいでいたのですが、ちょうどORFにlacZをノックインしたキメラマウスが理研から送られてきた頃で、そいつを使ってFISHをしたところ、6330403K07Rikのプロモーターから転写されたlacZのmRNAは細胞質に運ばれていたので、この魅力的な仮説もボツ。
目を奪われる美しい核内局在で華々しいデビューしたものの、その後の解析は、2021シーズンの後半戦の佐藤輝のごとく、打席に立てども立てども三振と凡打の嵐です。
しかし、逃げられれば逃げられるほど燃え上がる片思いの恋の如く、これはなにかものすごく重要なメカニズムが隠されているに違いない、と、なってしまうのも世の常です。KOマウスもできたし、ウエスタンできれいなシングルバンドが出る特異的なモノクローナル抗体もできたし、材料は揃った、あとは手を動かすだけだ、と、この核のシグナルが幻だとは露ほども思いませんでした。
でもこの仕事。これ以外にも、あまりにも奇妙なことが多すぎました。例えば、上の写真にもある通りNeuro2A細胞では6330403K07RikでFISHするととてもきれいな核内のシグナルが見えるのですが、ウエスタンでは全くバンドが見えません。細胞質に運ばれてしまう全長のmRNA発現ベクターを発現した場合はくっきりバンドが見えます。ここでまたおかしいな、と考えずに、ありそうにない仮説を積み重ねてしまいます。曰く、
- Neuro2Aの内在の6330403K07RikのmRNAは何らかのメカニズムが働いて核内に繋留されるために翻訳されない。
- Neuro2Aに発現させた外来のmRNAは細胞質に運ばれるので翻訳されて抗体でも検出できる。
- in vivoでは何らかのシグナルが入ってmRNAの核内繋留が解除されるために細胞質に輸送されて翻訳される。
- もしかしたらNeuro2A細胞でもいろいろな刺激を与えれば内在のmRNAが細胞質に輸送されて翻訳されるかも!!!
と、いうわけで、核内に繋留されたmRNAが輸送されて翻訳されるような条件を探すべく、Neuro2A細胞を水攻め熱攻め低酸素攻めNGF攻めといろいろこねくり回したのですが、FISHでの核内のmRNAのきれいなシグナルは一向に細胞質へと運ばれることもなく、内在mRNA由来のタンパク質が発現することもありませんでした。
そんなこんなで実験は完全に行き詰まり、にっちもさっちも行かなくなって、このプロジェクトは塩漬けフグの卵巣状態になってしまいました。この状況を打破してくれたのが、北大に移ってからラボにjoinしてくれた学生の高橋くんと講師の米田さんです。
#20の大迷走(3)に続く
投稿者プロフィール
最新の投稿
 ノンドメインブログ2025.06.27来し方行末~パート1
ノンドメインブログ2025.06.27来し方行末~パート1 ノンドメインブログ2025.02.11平和な日々
ノンドメインブログ2025.02.11平和な日々 ノンドメインブログ2024.11.25MBSJ2024のシンポジウムのすごいゲスト
ノンドメインブログ2024.11.25MBSJ2024のシンポジウムのすごいゲスト ノンドメインブログ2024.09.17奇遇癖
ノンドメインブログ2024.09.17奇遇癖